(一)DNA的二级结构双螺旋结构模型(double helixmodel)
1953年,Watson和Crick提出了著名的DNA分子的双螺旋结构模型,揭示了遗传信息是如何储存在DNA分子中,以及遗传性状何以在世代间得以保持。这是生物学发展的重大里程碑。
在DNA双螺旋结构模型建立之前,早在1868年,Miescher已经从脓细胞提取到核酸与蛋白质的复合物,当时称为核素(nuclein)。但核酸在生命活动中的重要地位,却迟至本世纪50年代才被认识。
本世纪20年代,Levene研究了核酸的化学结构并提出四核苷酸假说;40年代末,Avery,Hershey和Chase的实验严密地证实了DNA就是遗传物质;50年代初,Chargaff应用紫外分光光度法结合纸层析等简单技术,对多种生物DNA作碱基定量分析,发现DNA碱基组成有如下规律(表15-3)。
表15-3 不同生物来源的DNA四种碱基比例关系
| DNA来源 | 腺嘌呤(A) | 胸腺嘧啶(T) | 鸟嘌呤(G) | 胞嘧啶(C) | (A+T)/(G+C) |
| 大肠杆菌 | 25.4 | 24.8 | 24.1 | 25.7 | 1.01 |
| 小麦 | 26.8 | 28.0 | 23.2 | 22.7 | 1.21 |
| 鼠 | 29.7 | 25.6 | 21.9 | 22.8 | 1.21 |
| 猪:肝 | 29.4 | 29.7 | 20.5 | 20.5 | 1.43 |
| 胸腺 | 30.0 | 28.9 | 20.4 | 20.7 | |
| 脾 | 29.6 | 29.2 | 20.4 | 20.8 | |
| 酵母 | 31.3 | 32.9 | 18.7 | 17.5 | 1.079 |
(1)同一生物的不同组织的DNA碱基组成相同;
(2)一种生物DNA碱基组成不随生物体的年龄、营养状态或者环境变化而改变;
(3)几乎所有的DNA,无论种属来源如何,其腺嘌呤摩尔含量与胸腺嘧啶摩尔含量相同(A]=[T),鸟嘌呤摩尔含量与胞嘧啶摩尔含量相同(G]=[C),总的嘌呤摩尔含量与总的嘧啶摩尔含量相同([A+G]=[C]+[T)。
(4)不同生物来源的DNA碱基组成不同,表现在A+T/G+C比值的不同;
这些结果后来为DNA的双螺旋结构模型提供了一个有力的佐证。
Watson和Crick以立体化学原理为准则,对Wilkins和Franklin的DNa X射线衍射分析结果加以研究,提出了DNA结构的双螺旋模式,其主要内容如下:
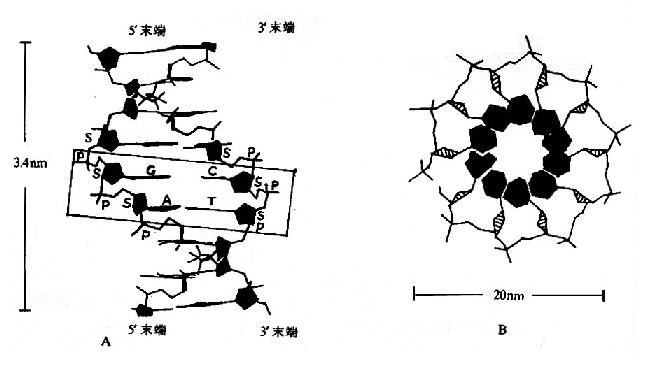
图15-5 DNA的双螺旋结构模式
A.正面观:长方框内有详细说明,S代表脱氧核糖。
B.俯视:涂黑的是碱基,此处全部碱基都是嘧啶,只看到糖的侧面略呈三角形,最外围是磷酸及其酯键。
(1)在DNA分子中,两股DNA链围绕一假想的共同轴心形成一右手螺旋结构,双螺旋的螺距为3.4nm,直径为2.0nm。(图15-5,A,B)。
(2)链的骨架(backbone)由交替出现的、亲水的脱氧核糖基和磷酸基构成,位于双螺旋的外侧。
(3)碱基位于双螺旋的内侧,两股链中的嘌呤和嘧啶碱基以其疏水的、近于平面的环形结构彼此密切相近,平面与双螺旋的长轴相垂直。一股链中的嘌呤碱基与另一股链中位于同一平面的嘧啶碱基之间以氢链相连,称为碱基互补配对或碱基配对(base pairing),碱基对层间的距离为0.34nm。碱基互补配对总是出现于腺嘌呤与胸腺嘧啶之间(A=T),形成两个氢键;或者出现于鸟嘌呤与胞嘧啶之间(G=C),形成三个氢键。(图15-6)。
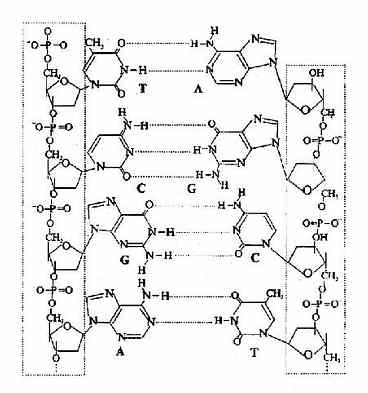
图15-6 A-T,G-C间的氢键形成
(4)DNA双螺旋中的两股链走向是反平行的,一股链是5′→3′走向,另一股链是3′→5′走向。两股链之间在空间上形成一条大沟(major groove)和一条小沟(minor groove),这是蛋白质识别DNA的碱基序列,与其发生相互作用的基础。
DNA双螺旋的稳定由互补碱基对之间的氢键和碱基对层间的堆积力(basestacking force)维系。DNA双螺旋中两股链中碱基互补的特点,逻辑地预示了DNA复制过程是先将DNA分子中的两股链分离开,然后以每一股链为模板(亲本),通过碱基互补原则合成相应的互补链(复本),形成两个完全相同的DNA分子。因为复制得到的每对链中只有一条是亲链,即保留了一半亲链,将这种复制方式称为DNA的半保留复制(semiconservativereplication)。后来证明,半保留复制是生物体遗传信息传递的最基本方式。
DNA双螺旋是核酸二级结构的重要形式。双螺旋结构理论支配了近代核酸结构功能的研究和发展,是生命科学发展史上的杰出贡献。
| 上一页:(三)限制性片段长度多态性 |
| 当前页:(一)DNA的二级结构双螺旋结构模型(double helixmodel) |
| 下一页:(二)DNA结构的多态性 |