(一)DNA的半保留复制(semiconservative replication)
Watson和Crick在提出DNA双螺旋结构模型时即推测,DNA在复制时首先两条链之间的氢键断裂两条链分开,然后以每一条链分别做模板各自合成一条新的DNA链,这样新合成的子代DNA分子中一条链来自亲代DNA,另一条链是新合成的,这种复制方式为半保留复制。
1958年Meselson和Stahl利用氮标记技术在大肠杆菌中首次证实了DNA的半保留复制,他们将大肠杆菌放在含有15N标记的NH4Cl培养基中繁殖了15代,使所有的大肠杆菌DNA被15N所标记,可以得到15N桪NA。然后将细菌转移到含有14N标记的NH4Cl培养基中进行培养,在培养不同代数时,收集细菌,裂介细胞,用氯化铯(CsCl)密度梯度离心法观察DNA所处的位置。由于15N桪NA的密度比普通DNA(14N-DNA)的密度大,在氯化铯密度梯度离心(density gradientcentrifugation)时,两种密度不同的DNA分布在不同的区带。
实验结果表明:在全部由15N标记的培养基中得到的15N桪NA显示为一条重密度带位于离心管的管底。当转入14N标记的培养基中繁殖后第一代,得到了一条中密度带,这是15N桪NA和14N-DNA的杂交分子。第二代有中密度带及低密度带两个区带,这表明它们分别为15N14N-DNA和14N14N-DNA。随着以后在14N培养基中培养代数的增加,低密度带增强,而中密度带逐渐减弱,离心结束后,从管底到管口,CsCl溶液密度分布从高到低形成密度梯度,不同重量的DNA分子就停留在与其相当的CsCl密度处,在紫外光下可以看到DNA分子形成的区带。为了证实第一代杂交分子确实是一半15N-DNA-半14N-DNA,将这种杂交分子经加热变性,对于变性前后的DNA分别进行CsCl密度梯度离心,结果变性前的杂交分子为一条中密度带,变性后则分为两条区带,即重密度带(15N-DNA)及低密度带(14N-DNA)。它们的实验只有用半保留复制的理论才能得到圆满的解释(图16-2和16-3)。
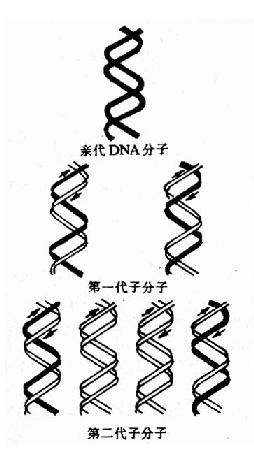 | 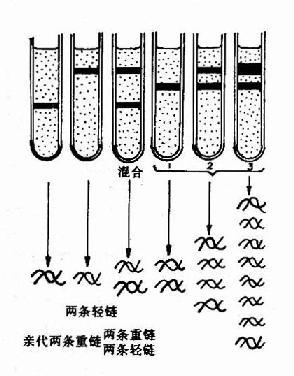 |
| 图16-2 DNA的半保留复制第一代分子含有一条亲代的链(用黑色素示),与另一条新合成的链(用白色表示)配对。在以后的连续复制过程中,原来亲代的两条链仍然保持完整,因此总有两个分子各具有一条原来亲代的链。 | 图16-3 DNA的半保留复制-MeslsonStahl实验密度梯度离心后的DNA位置:左三管为对照;右三管为实验结果 |
| 上一页:第一节 DNA的复制 |
| 当前页:(一)DNA的半保留复制(semiconservative replication) |
| 下一页:(二)DNA复制的一般过程: |