(һ)DNA�ľۺϷ�Ӧ��DNA�ۺ�ø
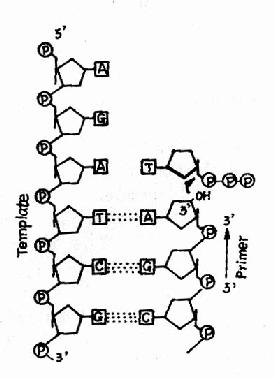
ͼ16��9��DNA�ۺ�ø������
1957�꣬Arthur kornberg�״��ڴ˾��з���DNA�ۺ�ø��(DNa polymerase ��,��дDNA pol��)��������̷�����DNA�ۺ�ø���DNA�ۺ�ø��(DNa polymerase ��дDNA pol��,DNA pol��)ʵ��֤���˾���DNA���Ƶ���Ҫ���̿�DNa pol�������ã���DNA pol���DNA pol����DNA�����У�������������á�����16��1��
����ø�Ĺ�ͬ�����ǣ�����ҪDNAģ�壬�������ø�ֳ�Ϊ����DNA��DNA�ۺ�ø(DNa dependent DNA polymerase, DDDP)������ҪRNA��DNA��Ϊ����(primer)����DNA�ۺ�ø���ܴ�ͷ��DNA����ʼ���۴�dNTP�ӵ������3��OHĩ�ˣ����DNA�ϳɵķ�����5���3�䡣ͼ16-9��������DNA�ۺ�ø�����ڶ��ø��������DNA���ƺ������̵IJ�ͬ�η������á�����DNA�ۺ�ø�����о�����������Ҵ���������DNA�ۺ�ø�Ļ����ص㣬�����������ؽ���DNa pol������ò�ָ���������DNA pol�������ԣ�
1.DNA�ۺ�ø��
DNA pol������һ����������ɣ�������Ϊ109KD��ø�����к���һ��Zn++���Ǿۺϻ��Ա���ġ�
�˾�ÿ��ϸ����Լ��400��ø���ӣ�ÿ��ø����ÿ������37�����ܴ�667����������뵽DNA���У��ÿݲݸ˾�����ø�ɽ���øˮ�������Ƭ�Σ���Ƭ�η�����Ϊ76KD��ͨ����ΪklenowƬ�Σ�СƬ��Ϊ34KD����СƬ�ξ��в�ͬ��ø���ԡ�
(1)DNA�ۺ�ø��5���3��ۺϻ��ԣ�
����DNA�ۺ�ø����Ҫ�Ļ��ԣ���ģ��DNA�ϵĺ�����˳��������dNTP����ӵ�����RNA3��OHĩ�ˣ����ٽ�3��OH��dNTP��5��PO4�γ������������ø��רһ�Ա���Ϊ�½����dNTP������ģ��DNA������ʱ���д����ã�5���3��ۺϻ��Դ�����klenowƬ����(ͼ16��9��ͼ16-10)��
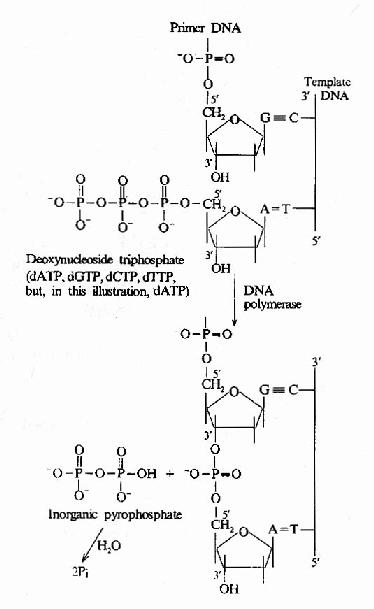
ͼ16-10��DNA�ۺ�ø����DNA���ӳ�
(2)DNA�ۺ�ø��3���5�����к���ø���ԣ�
����ø���Ե���Ҫ�����Ǵ�3���5�䷽��ʶ���г�DNA������ĩ����ģ��DNA����Զ�����ĺ����ᣬ���ֹ��ܳ�ΪУ�Թ��ܣ����DZ�֤��ۺ����õ���ȷ�Բ���ȱ�ٵģ���˶���DNA�����м��ߵı�������������Ҫ�ġ�
(3)DNA�ۺ�ø��5���3�����к���ø���ԣ�
����ø�����Ǵ�DNA����5�����3��ĩ��ˮ������Եĺ����ᣬ�������ж������������ÿ�����г�10�������ᡣ��ˣ�����ø������DNA���˵����п�������Ҫ���ã�����ɵ�DNAƬ��ȥ��5��˵�RNA����Ҳ�DZ���ġ�
DNA pol���5���3��ۺϻ��Ժ�5���3������ø����Эͬ���ã�����ʹDNAһ�����ϵ��пڴ�5���3�䷽���ƶ������ַ�Ӧ����ȱ��ƽ��(nick translation)�����ô˷�Ӧ���������DNAƬ�ν��з�������(��-32PdNTP)�ı���Ƴ�̽��(probe)�����к���ķ����ӽ�ʵ�飬���ִ���������ѧ��һ����Ҫ����(ͼ16��11)��
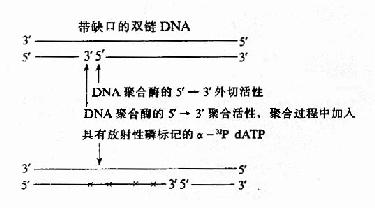
ͼ16-11��ȱ��ƽ�Ʊ��DNA̽��
����ʵ��֤ʵDNA pol����DNA���ƹ����е���Ҫø������������Ҫ��DNA���˺�����йء�
2.DNA�ۺ�ø��(DNA pol��)
��ø������Ϊ120KD��ÿ��ϸ��Լ��100��ø���ӣ�������ֻ��DNa pol���5%��������5���3��ۺϻ��Ժ�3���5�����л��ԣ���û��5���3�����л��ԣ��������ÿ�����DNA�������йء�
3.DNA�ۺ�ø��(DNA pol��)
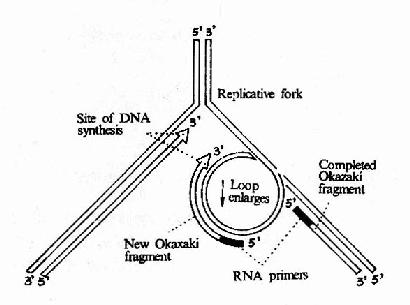
ͼ16��12��DNA�ۺ�ø����ȵ�������ӵĺϳ�
������DNA���ƹ���������Ҫ���õľۺ�ø��������һ�����ǻ���ɵĵ����ʷ��ӣ����������600kDa����ø�����γ�һ�����ԳƵĶ����壬ÿ���˾�ϸ����ֻ��10?0��ø���ӣ�����dNTP����DNA��������ȴ�����ģ�ԼΪ9000������/ÿ����/ÿ��ø���ӡ���Ҳ֤��DNa pol����DNA���ƹ�������Ҫ�������õ�ø���ڴ˾�Ⱦɫ��DNA���и���ʱ��DNA�ۺ�ø��ȫø�����ǵ��������õģ������������壬����ø�ȹ���һ��������(replisome)�����ڸ�����Ĵ��ڣ��ȵ��������������ͬʱ���ơ�DNa pol�����ɶ��ǻ���ɵIJ��Գƶ����壬������ͬʱ�����ȵ�����������ĸ��ƣ��ڦա�174�ĸ����й۲쵽���������ǰ�����DNA����(loop)�Ĵ��ڡ�ͼ16?2���Կ����������������ģ��DNA��DNA�ۺ�ø��ȫø����ת��180����γ�һ�����ۣ���˸���Ƭ�εĺϳɷ����ܹ����ȵ����ĺϳɷ����Լ��������ƶ�����һ�¡�
����DNA pol����ǰ�ƶ����ȵ����ĺϳ����ӳ���ͬʱ������Ƭ��Ҳ�ڲ����ӳ�����һ����Ҳ�ڲ�����������Ƭ�κϳɵ�ǰһ��Ƭ�ε�5���ʱ����һ�����۾��ͷų��������ڸ��Ʋ���ǰ�ƶ��ֿɽ���һ�����������ģ���û���������������ϳ��µ����Ȼ�����γ�һ��С�����ۣ������µĸ���Ƭ�εĺϳɡ��ɴ�ģ�Ͳ��ѿ�����������ĺϳ���Ҫ�����Ե������������ϳɽ���������ǰ�������һ������Ƭ�εij��ȡ�����Ƭ����ɺ���5��˵�RNA������DNa pol���5���3������ø�����г����ɴ���ɵĿ�϶����DNA pol���5���3��ۺϻ��Դ�dNTP�õ��������DNA�ĸ�������DNa pol���DNApol�����������ɵġ�
�����б�˵�����ִ˾�DNA�ۺ�ø������
��16��1���˾�DNA�ۺ�ø����
| DNA�ۺ�ø�� | DNA�ۺ�ø�� | DNA�ۺ�ø�� | |
| ������ | 109KD | 120KD | ��600KD |
| ÿ��ϸ���еķ����� | 400 | 17-100 | 10-20 |
| 5���3��ۺϻ��� | + | + | + |
| 37��ת���ʺ���������ø���ӡ����� | 600 | 30 | 30,000 |
| 5���3������� | + | - | - |
| 5���3������� | + | + | + |
| �п�ƽ�ƻ��� | + | - | - |
| ��dNTP���� | �� | �� | �� |
| ���� | �� | ���� | ���� |
| ȥ������ | |||
| ���ȱ |
�������DNA�ۺ�ø
�������DNA�ۺ�ø�Ц����¡��á��ļ��š����ǵĻ������������ڴ˾�DNA�ۺ�ø������Ҫ�����Ǵ�dNTP��5���3��ۺϻ��ԣ�������������16��2��
��16-2���������DNA�ۺ�ø
| �� | �� | �� | �� | �� | |
| �ǻ��� | 4 | 4 | 4 | 2 | 5 |
| ��������KD) | ��250 | 36-38 | 160-300 | 170 | 256 |
| ϸ���ڶ�λ | �� | �� | ������ | �� | �� |
| 5���3��ۺϻ��� | �� | �� | �� | �� | �� |
| 3���5������� | �� | �� | �� | �� | �� |
| ���� | ���ơ����� | �� | ���� | ���� | ���� |
���ϸ����DNA����������Ҫ���õ���DNA pol������Ҫ����Ⱦɫ��DNA�ĸ��ơ�DNa pol�µ�ģ���������Ǿ���ȱ�ڵ�DNA���ӣ�����Ϊ����DNA���йء�DNa pol����������DNA�ĸ����������á�DNA pol�IJ�����5���3��ۺϻ��ԣ����һ�����3���5������ø���ԣ�����Ϊ�������DNA��������DNa pol����DNA pol��Эͬ�����½��еģ�ǰ�����ĺϳɿ�DNA pol�Ĵ������һ���Ҫһ��ϸ�����ڵ������ӗ��ֳϸ���˿��?proliferatingcell nucleus antigen, PCNA)���롣��������ĺϳɿ�DNA pol��������ø���������ɡ�
| ��һҳ������DNA���Ƶ��ӳ����Լ������ø�͵����ʷ��ӣ� |
| ��ǰҳ��(һ)DNA�ľۺϷ�Ӧ��DNA�ۺ�ø |
| ��һҳ��(��)�볬�����ɳ��йص�ø�� |