(二)mRNA是合成蛋白质的直接模板
原核细胞中每种mRNA分子常带有多个功能相关蛋白质的编码信息,以一种多顺反子的形式排列,在翻译过程中可同时合成几种蛋白质,而真核细胞中,每种mRNA一般只带有一种蛋白质编码信息,是单顺反子的形式。mRNA以它分子中的核苷酸排列顺序携带从DNA传递来的遗传信息,作为蛋白质生物合成的直接模板,决定蛋白质分子中的氨基酸排列顺序。不同的蛋白质有各自不同的mRNA,mRNA除含有编码区外,两端还有非编码区。非编码区对于mRNA的模板活性是必需的,特别是5’端非编码区在蛋白质合成中被认为是与核糖体结合的部位。见图18-2。
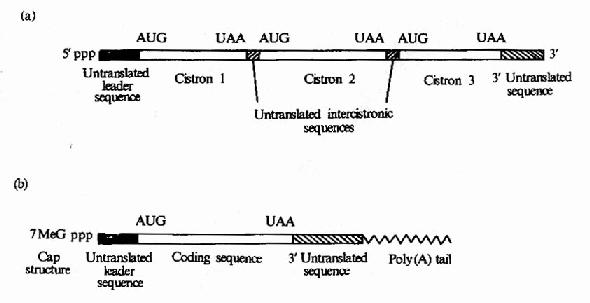
图18-2 (a)原核生物mRNA)为多顺反子(b)真核生物mRNA为单顺反子
mRNA分子上以5'→3'方向,从AUG开始每三个连续的核苷酸组成一个密码子,mRNA中的四种碱基可以组成64种密码子。这些密码不仅代表了20种氨基酸,还决定了翻译过程的起始与终止位置。每种氨基酸至少有一种密码子,最多的有6种密码子。从对遗传密码性质的推论到决定各个密码子的含义,进而全部阐明遗传密码,是科学上最杰出的成就之一,科学家们设计了十分出色的遗传学和生物化学实验,于1966年编排出了遗传密码字典。见表18-1。
表18-1 氨基酸的密码(code)
| 5’末端(第1位碱基) | 中间碱基(第二位碱基) | 3’末端(第三位碱基) | |||
| U | C | A | G | ||
| U | 苯丙(Pne)F | 丝(Ser)S | 酪(Tyr)Y | 半胱(Cys)C | U |
| 苯内(Pne) | 丝(Ser) | 酪(Tyr) | 半胱(Cys) | C | |
| 亮(Leu)L | 丝(Ser) | 终止信号 | 终止信号 | A | |
| 亮(Leu) | 丝(Ser) | 终止信号 | 色(Trp) | G | |
| C | 亮(Leu) | 脯(Pro)P | 组(His)H | 精(Arg)R | U |
| 亮(Leu) | 脯(Pro) | 组(His) | 精(Arg) | C | |
| 亮(Leu) | 脯(Pro) | 谷胺(Gin)Q | 精(Arg) | A | |
| 亮(Leu) | 脯(Pro) | 谷胺(Gin) | 精(Arg) | G | |
| A | |||||
| 异亮(ILe)I | 苏(Thr)T | 天胺(Asn)N | 丝(Ser)S | U | |
| 异亮(ILe) | 苏(Thr) | 天胺(Asn) | 丝(Ser) | C | |
| 异亮(ILe) | 苏(Thr) | 赖(Lys)K | 精(Arg)R | A | |
| *蛋(Met)M(起动信号) | 苏(Thr) | 赖(Lys) | 精(Arg) | G | |
| G | |||||
| 缬(Val)V | 丙(Ala)A | 天(Asp)D | 甘(Gly)G | U | |
| 缬(Val) | 丙(Ala) | 天(Asp) | 甘(Gly) | C | |
| 缬(Val) | 丙(Ala) | 谷(Glu)E | 甘(Gly) | A | |
| 缬(Val) | 丙(Ala) | 谷(Glu) | 甘(Gly) | G | |
*位于mRNA起动部位AUG为氨基酸合成肽链的起动信号。以哺乳动物为代表的真核生物,此密码子代表蛋氨酸;以微生物为代表的原核生物则代表甲酰蛋氨酸。
遗传密码具有以下几种特点:
(1)起始码与终止码(Initiation codon and termination codon):
密码子AUG是起始密码,代表合成肽链的第一个氨基酸的位置,它们位于mRNA5′末端,同时它也是蛋氨酸的密码子,因此原核生物和真核生物多肽链合成的第一个氨基酸都是蛋氨酸,当然少数细菌中也用GUG做为起始码。在真核生物CUG偶尔也用作起始蛋氨酸的密码。密码子UAA,UAG,UGA是肽链成的终止密码,不代表任何氨基酸,它们单独或共同存在于mRNA3’末端。因此翻译是沿着mRNA分子5′→3′方向进行的。
(2)密码无标点符号:两个密码子之间没有任何核苷酸隔天,因此从起始码AUG开始,三个碱基代有一个氨基酸,这就构成了一个连续不断的读框,直至终止码。如果在读框中间插入或缺失一个碱基就会造成移码突变,引起突变位点下游氨基排列的错误。
(3)密码的简并性(Degemeracy):
一种氨基酸有几组密码子,或者几组密码子代表一种氨基酸的现象称为密码子的简并性,这种简并性主要是由于密码子的第三个碱基发生摆动现象形成的,也就是说密码子的专一性主要由前两个碱基决定,即使第三个碱基发生突变也能翻译出正确的氨基酸,这对于保证物种的稳定性有一定意义。如:GCU,GCC,GCA,GCG都代表丙氨酸。
(4)密码的通用性:
大量的事实证明生命世界从低等到高等,都使用一套密码,也就是说遗传密码在很长的进化时期中保持不变。因此这张密码表是生物界通用的。然而,出乎人们预料的是,真核生物线粒体的密码子有许多不同于通用密码,例如人线粒体中,UGA不是终止码,而是色氨酸的密码子,AGA,AGG不是精氨酸的密码子,而是终止密码子,加上通用密码中的UAA和UAG,线粒体中共有四组终止码。内部甲硫氨酸密码子有两个,即AUG和AUA;而起始甲硫氨酸密码子有四组,即AUN。
密码子结构与氨基酸侧链析性之间也有一定关系。①氨基酸侧链极性性质在多数情况下由断面子的第二个碱基决定。第二个碱基为嘧啶(Y)时,氨基酸侧链为非极性,第二个碱基为嘌呤时,氨基酸侧链则有极性。②当第一个碱基为U或A,第二个碱基为C,第三个碱基无特异性时,所决定的氨基酸侧链为极性不带电。③当第一个碱基不是U,第二个碱基是P时,氨基酸侧链则带电。在此前提下,若是一个是C或A时,表示带正电的氨基酸,第一、二个碱基分别是G、A时,此种氨革酸带负电,但上述关系也有个别例外。
一种氨基酸由多种密码子所编码的事实使人想到:同一种氨基酸的一组密码子的使用频率是否相同?许多实验证实,在原核生物和高等真核生物中同一组密码子的使用频率是不相同的。高频密码子多出现在那些表达量高的蛋白质基因中,例如,核糖体蛋白质基因,RecA蛋白质基因等。表18-2。这种使用频率与细胞内一组tRNA中的不同tRNA含量有关。
| 上一页:(一)合成原料 |
| 当前页:(二)mRNA是合成蛋白质的直接模板 |
| 下一页:(三)tRNA是氨基酸的运载工具: |