(��)��ʽ��������(trans��actingfactors)
�Է�ʽ����Ӱ��ת¼�����ӿ�ͳ��Ϊת¼����(transcription factors, TF)��RNA�ۺ�ø��һ�ַ�ʽ������ת¼�ĵ������ӡ������ϸ����RNA�ۺ�øͨ�����ܵ�������ת¼���ã�����Ҫ������ת¼���ӹ�ͬЭ������RNA�ۺ�ø����Ӧ��ת¼���ӷֱ��ΪTF��TF��TF��TF���о���ࡣ��19-2�г���˻���ת¼��Ҫ������TF��
��19-2��RNA�ۺ�ø��Ļ���ת¼����
| ת¼���� | ��������kD) | ���� |
| TBP | 30 | ��TATA�� |
| TF��-B | 33 | �鵼RNA�ۺ�ø��Ľ�� |
| TF��-F | 30,74 | ����ø |
| TF��-E | 34,37 | ATPø |
| TF��-H | 62,89 | ����ø |
| TF��-A | 12,19,35 | �ȶ�TF��-D�Ľ�� |
| TF��-I | 120 | �ٽ�TF��-D�Ľ�� |
��ǰ��Ϊ��TATA�н�ϵĵ���������TF��D����������TF��Dʵ�ʰ�������ɷ֣���TATA�н�ϵĵ�����TBP(TATAbox binding protein)����Ψһ��ʶ��TATA�в������ϵ�ת¼���ӣ�������RNA�ۺ�øת¼ʱ����Ҫ�ģ�������ΪTBP�������(TBP��associated factors TAF)�����ٰ���8������TBP���ܽ�ϵ����ӡ�ת¼ǰ����TF��D��TATA�н�ϣ��̶�TF��B����C����TBP��DNA�������ϣ���N��������RNA�ۺ�ø���ͽ�ϣ������������ǻ���ɵ�TF��F����װ�䣬TF��F����RNA�ۺ�ø�γɸ����壬������������ATP����������DNA����ø���ԣ��ܽǰ����DNA˫��������ת¼�������������á����������������о���TF��D��B��F��RNA�ۺ�ø�����γ�һ��������ȡ�����ת¼���ܻ�����ת¼ǰ��ʼ������(pre��intitiationcomplex, PIC)����ת¼mRNA��TF��H�Ƕ��ǻ��������壬����������ATP����������DNA����ø���ԣ���ת¼�������з������ã�TF��E�������ǻ���ɵ��ľ��壬��ֱ����DNA��϶���������TF��B��ϵ�������ATPø�Ļ��ԣ�TF��E��TF��H�ļ�����γ�������ת¼������(ͼ19?5)����ת¼�������ɳ���RNA��TF��A���ȶ�TF��D��TATA�еĽ�ϣ����ת¼Ч�ʣ�������ת¼������һ����Ҫ�ġ�
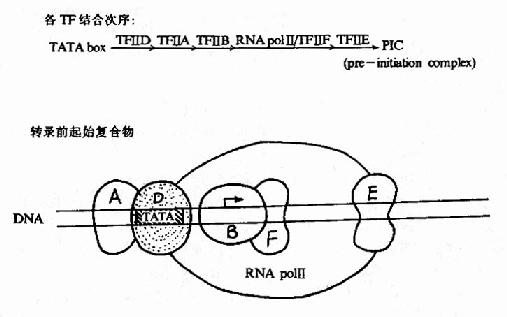
ͼ19-15��RNA�ۺ�ø��ת¼��������γ�ʾ��ͼ
���������ǵ��͵���������ת¼��������γɣ����е���������Ӳ���TATA�л�ͨ��TATA�п�ʼת¼�������е���TATA�е��������ǿ�TF��I��TF��D��ͬ����ȶ���ת¼��ʼ�����忪ʼת¼�ġ��ɴ˿��Կ������ת¼��ʼ�ĸ����ԡ�
��ͬ�����ɲ�ͬ������������Ԫ����ɣ����벻ͬ��ת¼���ӽ�ϣ���Щת¼����ͨ���������ת¼���������ö�Ӱ��ת¼��Ч�ʡ������Ѿ����������ͬ��ת¼���ӣ������������ǣ�ͬһDNA���пɱ���ͬ�ĵ���������ʶ����ֱ�ӽ��DNA���еĵ�������������������ͬ�ĵ������Ӽ��������ã��������ת¼������ͨ�������ʣ������ʼ�������DNA������ϵ��Ӱ��ת¼Ч�ʵ�(��ͼ19��16)��ת¼����֮���ת¼������DNA�Ľ�϶���������ı仯���Ӷ�Ӱ��ת¼��Ч�ʡ�
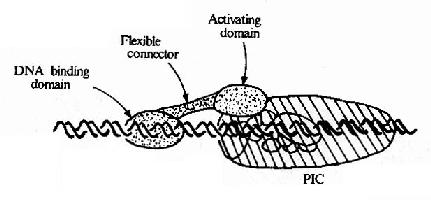
ͼ19-16��ת¼������ת¼�����������ģʽͼ
ͼ19-��6��ʾ����Ϊ�����ʵ�ת¼���Ӵӹ����Ϸ�����ṹ�ɰ����в�ͬ����DNA�����(DNa binding domain)������60��100��������л���֯�ļ���������ɣ���ת¼������(activating domain)������30��100������л���ɣ���ṹ���и����������ᡢ�����Ȱ�����������������Ȳ�ͬ���࣬�����Խṹ�������������������������������ṹ��IJ��֡�����DNAֱ�ӽ�ϵ�ת¼����û��DNA�������ͨ��ת¼������ֱ�ӻ���������ת¼�������Ӱ��ת¼Ч�ʡ�
��DNA��ϵ�ת¼���Ӵ���Զ�������ʽ�����ã���DNA��ϵĹ����������¼��֣�
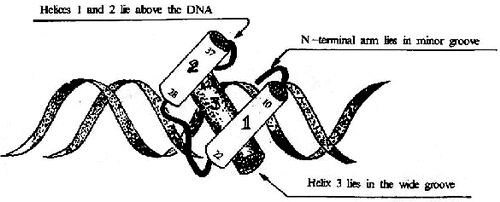
ͼ19��17��HTH�ṹ������DNA�Ľ��
��������ת�Ǫ�����(helix��turn��helix, HTH)������-��-����(helix��loop��helix,HLH)������ṹ����������������������ɶ��Ķ��γɵ�ת�ǻ����ӣ�����������motif�ṹ�Զ�������ʽ���������������൱��DNAһ���ݾ�(3.4nm)�������������պ÷ֱ�Ƕ��DNA���(ͼ19��17)��
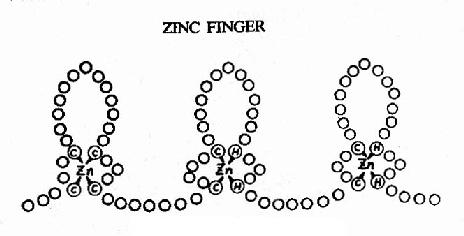
ͼ19��18�������ʵ�пָ�ṹ
��пָ(zinc finger)����ṹ��ͼ19��18��ʾ��ÿ���ظ��ġ�ָ��״�ṹԼ��23��������л���п��4����ۼ���4�����װ��ᡢ��2�����װ����2���鰱�����ϡ����������ʷ��ӿ���2?��������пָ�ظ���λ��ÿһ����λ������ָ������DNA˫����������Ӵ�5�������ᡣ������GC�н�ϵ�ת¼����SP1�о���������3��пָ�ظ��ṹ��
�ۼ��ԣ�����������(basic leucine zipper, bZIP)���ýṹ���ص��ǵ����ʷ��ӵ�������ÿ��6�����������һ��������л�������͵�����Щ������л����ڦ�������ͬһ��������֡�������ͬ�ṹ������������л���������ˮ����ϳɶ����壬�ö��������һ�˵��Ķθ�����������л��������������DNA˫�������ϴ�����ɵ�������Ž�ϡ������γɶ��������DNA���ͽ�������Խ��͡��ڸ��ࡢС����Ƥ��֬��ϸ����ijЩ��ϸ�����г�ΪC/EBP�����һ���൰�����ܹ���CAAT�кͲ�����ǿ�ӽ�ϣ��������������γ�bZIP������ṹ��
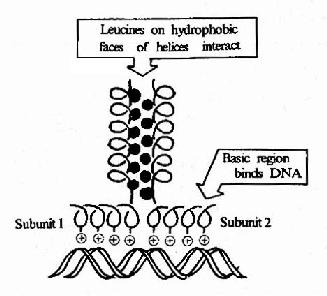
ͼ19��19�����������������ṹ������DNA�Ľ��
�������ɼ���ת¼���ص�ʵ�����ڵ�������DNA���������뵰����֮�������ã�����ı仯���ǵ����ʺͺ��ᡰ��ı��֡������������Ӽ�ı��ϡ�����á��ṹ�ϵı仯������������е����壬���ǵ���ʶ���о���ֻ���Σ�������������������Ҫ�Ĺ������ǿ�������һ����֪���д���Ŭ��̽����
| ��һҳ��(һ)˳ʽ�������(cis��acting elements) |
| ��ǰҳ��(��)��ʽ��������(trans��actingfactors) |
| ��һҳ��������Ҫ |