三、抑癌基因
抑癌基因又称肿瘤抑制基因,它的发现较癌基因晚,迄今克隆到的抑癌基因的数目亦较少,这并不意味着客观存在的抑癌基因就一定比癌基因少,只是由于技术上的原因,要想分离、鉴定、确认一个抑癌基因比较困难。
早在六十年代,有人将癌细胞与同种正常双倍体成纤维细胞融合,所获杂种细胞的后代只要保留某些正常亲本染色体时就可表现为正常表型。然而,随着染色体的丢失又可重新出现恶变细胞。这一现象表明,正常染色体内可能存在某些抑制肿瘤发生的基因,它们的丢失、突变或失去功能,好可使潜在的致癌因素如激活的癌基因发挥作用而致癌。
遗传学分析表明,人类的许多肿瘤细胞都有隐性遗传损害,已在染色体上定位的损害见表22-5。
表22-5 人类肿瘤的隐性遗传损害
| 肿 瘤 | 受损害的染色体 |
| 神经母细胞瘤、黑色素瘤、甲状腺癌嗜铬细胞瘤、MEN2 | 1p |
| 乳癌 | 1p |
| 小细胞肺癌、宫颈癌、肾细胞癌、肺腺癌 | 3p |
| 结直肠癌、家族性息肉 | 5p |
| 膀胱癌 | 9p |
| 星状细胞瘤、MEN2 | 10p |
| 膀胱癌、乳癌、横纹肌肉瘤、肝母细胞瘤胚胎瘤、肾母细胞瘤(Wilm's Tumor)肺癌 | 11p |
| MEN1 | 11p |
| 视网膜母细胞瘤、成骨肉瘤、小细胞肺癌、胃癌、结肠癌、乳癌 | 13p |
| 小细胞肺癌、结肠癌、乳癌、成骨肉瘤、星状细胞瘤、肺磷癌 | 17p |
| NF1 | 17p |
| 结肠癌 | 18p |
| 听神经瘤、脑膜瘤、NF2、嗜铬细胞瘤 | 22p |
抑癌基因概念是在研究视网膜母细胞瘤(Retinoblastoms ,RB)的遗传损害时提出来的。RB有家族性和散发性两种类型,其发闰机制不同。前者有先天的隐性遗传损害,其种系基因是有缺陷的,患RB的频率可高达80-90%,且往往是双侧,散发性RB,两次体细胞突变发生在同一个细胞,机率很小,患病也是单侧,Kundson早在1971年就提出RB发病的“两次击中学说”。现代分子遗传学分析手段的发展充分支持这一学说(图22-5)。1986年Draper统计,RB携带者发生第二原发癌的机率比一般人群要高数百倍。
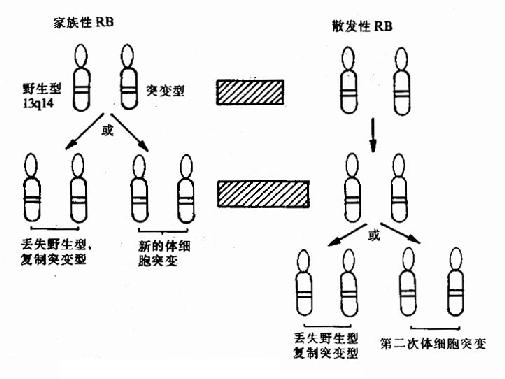
图22-5
关于抑癌基因如何起作用所知甚少,总体上总对生长起着控制作用,是一类生长控制基因或负调控基因,若功能丧失则失去负调控,细胞只能接受正调控信号,抑癌基因产物的功能多种多样,已确定的几中抑癌基因产物及其功能如表22-6。
表22-6 已确定的几种抑癌基因
| 基因 | 染色体定位 | 相关肿瘤 | 基因产物及功能 |
| RB | 13q14 | RB、成骨肉瘤、胃癌、SCLC、乳癌、结肠癌 | p105,控制生长 |
| WT | 11p13 | WT、横纹肌肉瘤、肺癌、膀胱癌、乳癌、肝母细胞瘤 | WT-ZFP,负调控转录因子 |
| NF-1 | 17p12 | 神经纤维瘤、嗜铬细胞瘤、雪旺氏细胞瘤、神经纤维肉瘤 | GAP,拮抗p21rasB |
| DCC | 18q21.3 | 结肠瘤 | P192, 细胞粘附分子 |
| p538 | 17p13 | 星状细胞瘤、胶质母细胞瘤、结肠癌、乳癌、成骨肉瘤、SCLC、胃癌、磷状细胞肺癌 | P53 控制生长 |
| erb A | 17q21 | ANLL | T3受体,含锌指结构的转录因子 |
| 上一页:二、细胞癌基因的激活 |
| 当前页:三、抑癌基因 |
| 下一页:提要 |