(三)远球小管和集合管
在远曲小管和集合管,重吸收大约12%滤过的Na+和CI-,分泌不同量的K+和H+,重吸收不同量的水。水、NaCI的重吸收以及K+和H+的分泌可根据机体贴的水、盐平衡状况来进行调节。如相机缺水或缺盐时,远曲小管和集合管可增加水、盐的重吸收;当机体水、盐过剩时,则水、盐重吸收明显减少,水和盐从尿排出增加。因此,远曲小管和集合管对水和盐的转运是可被调节的。水的重吸收主要受抗利尿激素调节,而Na+和K+的转运主要受醛固酮调节。
远曲小管和集合管上皮细胞间隙的紧密连接对小离子如Na+、K+和CI-等的通透性低,这些离子不易通过紧密连接回漏至小管腔内,因此,所能建立起来的管内外离子浓度梯度和电位梯度大。在远曲小管初段,对水的通透性很低,但仍主动重吸收NaCI,继续产生低渗小管液。Na+在远曲小和集合管的重吸收是逆较大的电化学梯度进行的,是主动重吸收过程。这可能与远曲小管的Na+泵在肾单位中活性最高有关。有人认为在远曲小管初段的小管液中,Na+是通过Na+-CI-同向转运进入细胞的,然后由Na+泵将Na+泵出细胞而主动重吸收回血(图8-13A)。NaCI同向转运体可被噻嗪类利尿药所抑制。
远曲小管后段和集合管含有两类细胞,即主细胞和闰细胞。主细胞重吸收Na+和水,分泌K+,闰细胞则主要分泌H+,主细胞重吸收Na+主要通过管腔膜上的Na+通道。管腔内的Na+顺电化学梯度通过管膜上的Na+通道进入细胞,然后,由Na+泵泵至细胞间液而被重吸收(图8-13B)。
K+的分泌 尿中K+的排泄量视K+的摄入量而定,高钾饮食可排出大量的钾,低钾饮食则尿中排钾量少,使机体的钾摄入量与排出量保持平衡,维持机体K+浓度的相对恒定。
K+分泌的动力包括:①在远曲小管和集合管的小管液中,Na+通过主细胞的管腔膜上的Na+通道进入细胞,然后,由基侧膜上的Na+泵将细胞内的Na+泵至细胞间隙而被重吸收,因而是生电性的,使管腔内带负电位(-10?40mV)。这种电位梯度也成为K+从细胞分泌至管腔的动力;②在远曲小管后段和集合管的主细胞内的K+浓度明显高于小管液中的K+浓度,K+便顺浓度梯度从细胞内通过管腔膜上的K+通道进入小管液;③Na+进入主细胞后,可刺激基侧膜上的Na+,使更多的K+从细胞外液中泵入细胞内,提高细胞内K+浓度,增加细胞内和小管液之间的K+浓度梯度,从而促进K+分泌,因此,K+的分泌与Na+的重吸收有密切关系。(图8-13B)。
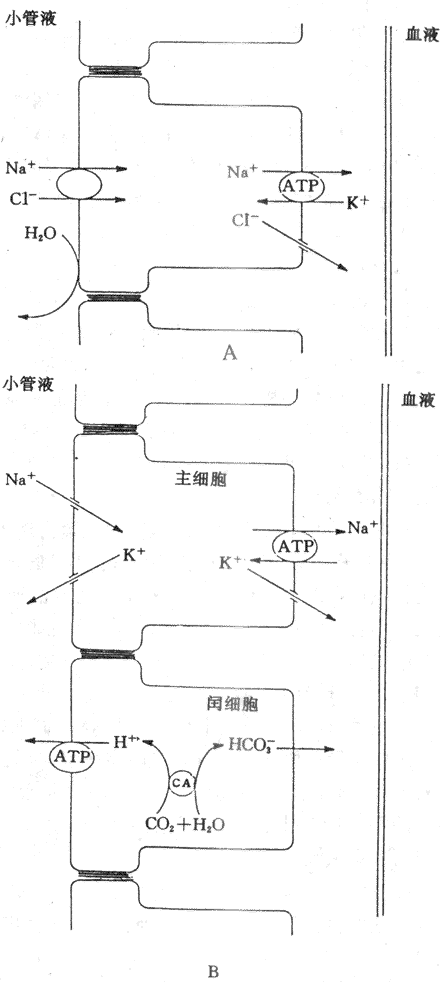
图8-13 远球小管和集合管重吸收NaCI、分泌K+和H+的示意图
A:远曲小管初段 B:远曲小管后段和集合管
H+的分泌除了近球小管细胞通过Na+-H+交换分泌H+,促进NaHCO3重吸收外,远曲小管和集合管的闰细胞也可分泌H+。H+的分泌是一个逆电化学梯度进行的主动转运过程。有人认为管腔膜上有H+泵,能将细胞内的H+历史意义入小管腔内。细胞内的CO2和H2O在碳酸酐酶催化作用下生成的H+和HCO3,H+由H+泵泵至小管液,HCO3则通过基侧膜回到血液中,因而H+分泌和HCO3的重吸收与酸碱平衡的调节有关(图8-13B)。闰细胞分泌的H+与HPO4-2结合形成H2PO4,这是可滴定酸;分泌的H+可与上皮细胞分泌的NH3结合,形成NH4+。可滴定酸和NH4+都不易透过管腔膜进入细胞而留在小管液中。因此,它们是尿液酸碱度的决定因素。
NH3的分泌远曲小管和集合管的上皮细胞在代谢过程中不断地生成NH3,这些NH3主要由谷氨酰胺脱氨而来。NH3具有脂溶性,能通过细胞膜向小管液周围组织间液和小管液自由扩散,扩散量取决于两种液体的pH值。小管液的pH较低(H+浓度较高),所以NH3较易向小管液中扩散。分泌的NH3能与小管液中的H+结合并生成NH4-,小管液中NH3浓度因而下降,于是管腔膜两侧形成了NH3浓度梯度,此浓度梯度又加速了NH3向小管液中扩散。由此可见,NH3的分泌与H+的分泌密切相关;H+分泌增加促使NH3分泌增多。NH3与H+结合并生成NH4-后,可进一步与小管液中的强酸盐(如NaCI等)的负离子结合,生成酸性铵盐(NH4CI等)并随尿排出。强酸盐的正离子(如Na+)则与H+交换而进入肾小管细胞,然后和细胞内HCO3-一起被转运回血。所以,肾小管细胞分泌NH3,不仅由于铵盐形成而促进了排H+,而且也促进了NaHCO3的重吸收。
| 上一页:(二)髓袢 |
| 当前页:(三)远球小管和集合管 |
| 下一页:第四节 尿液的浓缩和稀释 |