(三)耳蜗的生物现象
在耳蜗结构中除了能记录到与听神经纤维兴奋有关的动作电位,还能记录到一些其他形式的电变化。在耳蜗未受到刺激时,如果把一个电极放在鼓阶外淋巴中,并接地使之保持在零电位,那么用另一个测量电极可测出蜗管内淋巴中的电位为+80mV左右,这称为内淋巴电位。如果将此测量电极刺入毛细胞膜内,则膜内电位为-70?/FONT>-80mV。毛细胞顶端膜外的浸浴液为内淋巴,则该处毛细胞内(相当于-80mV)和膜外(相当于+80mV)的电位差当为160mV;而在毛细胞周围的浸浴液为外淋巴(电位相当于零),该处膜内外的电位差只有80mV左右;这是毛细胞静息电位和一般细胞不同之处。据实验分析,内淋巴中正电位的产生和维持,同蜗管外侧壁处的血管纹结构的细胞活动有直接关系(图9-16),并且对缺 O2非常敏感;有人发现,血管纹细胞的膜含有大量活性很高的ATP酶,具有“钠泵”的作用,它们可依靠分解ATP获得能量,将血浆中的K+泵入内淋巴,将内淋巴中的Na+泵入血浆,但被转运的K+担超过了Na+的量,这就使内淋巴中有大量K+蓄积,因而使内淋巴保持了较高的正电位;缺O2使ATP的生成受阻,也使Na+泵的活动受阻,因而使内淋巴的正电位不能维持。
当耳蜗接受声音刺激时,在耳蜗及其附近结构又可记录到一种特殊的电波动,称为微音器电位。这是一种交流性质的电变化,在一定的刺激强度范围内,它的频率和幅度与声波振动完全一致(图9-19);这一现象正如向一个电话机的受话器或微音器(即麦克风)发声时,它们可将声音振动转变为波形类似的音频电信号一样,这正是把耳蜗的这种电变化称为微音器电位的原因。事实上,如果对着一个实验动物和耳廓讲话,同时在耳蜗引导它的微音器电位,并将此电位经放大后连接到一个扬声器,那么扬声器发出的声音正好是讲话的声音!这一实验生动地说明,耳蜗在这里起着类似微音器的作用,能把声波变成相应的音频电信号。微音器电位的其它一些特点是:潜伏期极短,小于0.1ms;没有不应期;对缺O2和深麻醉相对地不敏感,以及它在听神经纤维变性时仍能出现等。
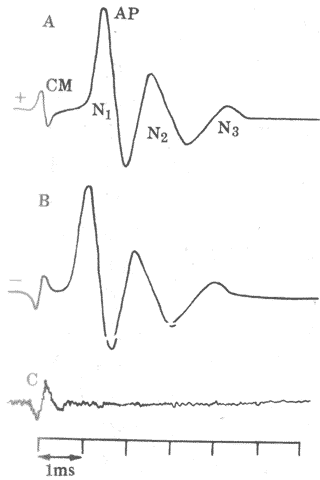
图9-19 由短声刺激引起的微音器电位和听神经动作电位
CM:微音器电位 AP:耳蜗神经动作电位(包括N1、N2、N3三个负电位)
A与B对比表明,声音位相改变时,微音器电位位相倒转,但神经动作
电位位相没有变化 C:在白噪音作用下,AP消失,CM仍存在
用微电极进入毛细胞的细胞内电变化记录的实验证明,所谓微音器电位就是多个毛细胞在接受声音刺激时产生的感受器电位的复合表现;在记录单一毛细胞跨膜电位的情况下,发现听毛只要有0.1。的角位移,就可引起毛细胞出现感受器电位,而且电位变化的方向与听毛受力的方向有关,亦即此电位既可是去极化的;这就说明了为什么微音器电位的波动同声波振动的频率和幅度相一致。
由于听毛的角位移和产生感受器电位之间只有一极短潜伏期,因而认为后者的产生是由于毛细胞顶部膜中有机械门控通道的存在,听毛受力引起该处膜的轻微变形,就足以改变这种通道蛋白质的功能状态,引起跨膜离子移动和相应的电位反应。在毛细胞,它的感受器电位可引起细胞基底部的递质(可能是谷氨酸和门冬氨酸)释放量的改变,进而引起分布在附近的耳蜗传入纤维产生动作电位,传向听觉高级枢,产生听觉。至于内毛细胞和外毛细胞在功能上有何不同,有人首先注意到它们所接受的传入纤维的数目有极大差异。据计算,人一侧耳蜗内毛细胞的总数约为3500个,外毛细胞则有约15000个,但来自螺旋神经节的约32000条听神经传入纤维中约有90%分布到内毛细胞的底部,这说明一个内毛细胞可接受多条传入纤维的分布,而多个外毛细胞才能接受一个传入纤维的轴突分支。因此一般认为,内毛细胞的作用是把不同频率的声音振动转变为大量分布在它们底部的传入纤维的神经冲动,向中枢传送听觉信息,而息细胞的作用近年来却发现有些特殊。有人发现毛细胞在基底膜振动和听毛受力而出现微音器电位时,此细胞可产生形体长短的快速改变,超极化引起细胞伸长,去极化引起细胞缩短,它们的形体改变因此也和外来声音振动的频率和振幅同步。据认为,外毛细胞的这种形体改变可以使所在基底膜部分原有的振动增强,亦却对行经该处的行波起放大作用,这显然使位于该部分基底膜上的内毛细胞更易受到刺激,提高了对该振动频率的敏感性。外毛细胞因膜内外电位差改变引起的机制尚不清楚,但这使得基底膜不仅仅是以固定的结构“被动”地对外界的振动产生行波,它还可以“主动”地增强行波的振动幅度。
| 上一页:(二)基底膜的振动和行波理论 |
| 当前页:(三)耳蜗的生物现象 |
| 下一页:四、听神经动作电位 |