�ġ���ά����ԭת��Ϊ��ά����
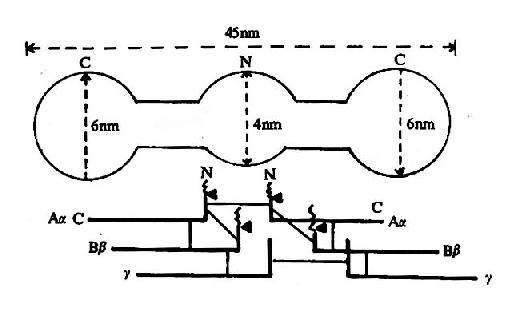
ͼ10��8����ά�ص���ԭ����ʾ��ͼ
�ϰ�Ϊ�羵�µķ�����״���°�ʾ6����������
һΪ˫�������Ϊ��Ѫø���õ�
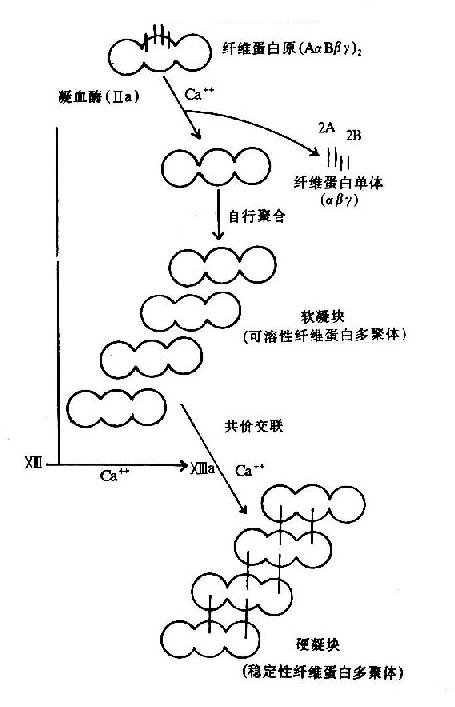
ͼ10��9����ά�ص�������������
ѪҺ���̵�ʵ������ά�������������ɣ�����Ѫ������ά����ԭ(fibrinogen)����Ѫø�����½���Ϊ��ά���ײ��ۺϳɲ����Ե���״�ṹ��
��ά����ԭ���������Ԧ��������£������ã�����ɣ�ÿ3������(�����¡���)�ʺϳ���״���γ�������״��������Nĩ���ж����ʹ̬�����ӵõ��ȶ���������������N���˷ֱ���һ��16����14���������С�ģ���Ϊ��ά��A��B����ˣ���ά����ԭ��дΪ(A��B�¦�)2(ͼ10-8)��
��Ѫø�ı���Ϊһ�ֵ���ˮ��ø����������������A����B�����ϵľ������ļ����г�A��B��ά�ġ�����ά��A��B��Ϊ�����ģ����϶ฺ��ɡ����ڵ���ų������谭��ά����ԭ֮��ۺϡ��г���ά��A��Bת��Ϊ��ά�����Լ�С��ͬʱ��¶�˻������λ�㣬�������Զ��ۺϣ���ά����λͨ���߿��ߡ��˿��˵ľۺ��γɾۺ��������ֶ���岻�ȶ�����Ϊ������(soft clot)������ͨ������X��a�����ý���ι̵���������X��aΪת��ø���ܴ�һ������ĹȰ���л��Ħã��Ȼ�����һ�����������л��İ���֮���γɹ��۽�ϣ�����ͳ�NH3(ͼ10��9��10)����ˣ�����X��a��Ϊ��ά�����ȶ�����(fibrin stabilizingfactor,FSF)������X�������ѪС�弰Ѫ���У�����Ѫø�г������Ķκ���ΪX��a��
�ɴ˲������ȶ���ά�������������鲻ͬ��������5M���弰1%��������Һ�в��ܽ⡣��ѪС���Ѫ˨�������������£�����״�ṹ�����������˿ڱ�Ե�˴˿����������˿ڱպϡ�����άϸ���ı������һ��������ά���ĵ����ʣ���ճ�����ף����ɢ�a������ά���������������ά���̶����������ԣ����Ӣ�a��ֱ�Ӳ����˿ڵ����ϡ�
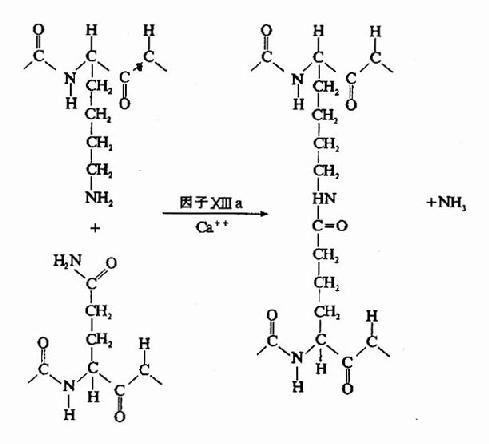
ͼ10��10������X��a���û���
�ܽ�������Ѫ���̿ɹ��ɳ������ص㣺
1.��Ѫ���ӵĻ������Ϊ�����ʵ�����ˮ�⣬��������Ѫ���˱������ǵ���ø������ø���õĵ����Щ����Ϊ����ø����Ѫ����(��������������)�İ�����˳������ƣ�������˿���ᵰ��øͬԴ���������ĵ�˿����л������ļ���ˮ�⡣C����Լ250��������л�ͬԴ�Ժܸߣ��Ǿ��д����ԵĽṹ��N���˵İ��������б仯�ϴ�������Ѫ�������õ����רһ�ԡ����Ǵ��ķ�Ӧ��Ca��������֬�μӡ�
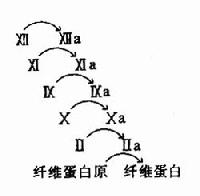
ͼ10��11��ѪҺ���̵��ٲ�ЧӦ
2.��֬����(��Դ��;����ѪС�壬��Դ��;������֯��Ѫ�����ṩ)ʹ���Ӧ�ڽ���������У������߷�Ӧ�ٶȣ���Ca�������������ڴٽ�ø�͵�������֬����Ľ�ϡ�
3.��Ѫ���ӻ���ٲ�ЧӦ(cascade)ʹѪҺ���̾��и�Ч�ʺ;��ܵ��ص���������ͼ10��11��ʾ��
4.ά����K���ڡ���Դ����Ѫ�о�����Ҫ���á�
5.��Ѫ�����е�������ʹ��Ӧ���ϼ��٣����ղ�����ά�����п���Ѫ���á���������Ѫ�뿹��Ѫ��������ϵ�ġ�
| ��һҳ��������Ѫøԭ�ļ��� |
| ��ǰҳ���ġ���ά����ԭת��Ϊ��ά���� |
| ��һҳ���塢��Ѫ���õĵ��� |